





بطور کلی در سیستم های یوکاریوتی، تغییرات هیستون، ساختار کروماتین را برای رونویسی از طریق تشکیل هتروکروماتین ها متوقف می کند. تقریبا در همه موجودات، تشکیل هتروکروماتین نیازمند آن است که هیستون H3 کروماتین، استیل زدایی (داستیله) شده و سپس در لیزین ۹ متیله شود.
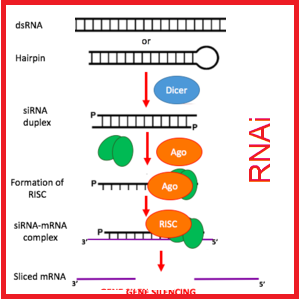
گاهی اوقات در حالت طبیعی، یک رشته از منطقه سانترومری بیان می شود، در حالی که رشته مکمل آن که در معرض خاموشی حاصل از هتروکروماتینه شدن قرار دارد، رونویسی می شود. چنین نسخه برداری منجر به تشکیل dsRNA می شود که بوسیله یک مرحله پردازش که در درون هسته واقع شده و مشابه با RNAi می باشد، انجام می شود، در ادامه siRNA تشکیل شده سبب فعال شدن کمپلکسی می شود که حاوی آنزیم متیل ترانسفر از هیستون است. چنین کمپلکسی منجر به جفت شدن بازهای DNA – RNA می شود و هیستون H3 – K9 این منطقه ممکن است متیله شود که در نهایت منجر به تشکیل هتروکروماتین در ناحیه هدف می گردد.
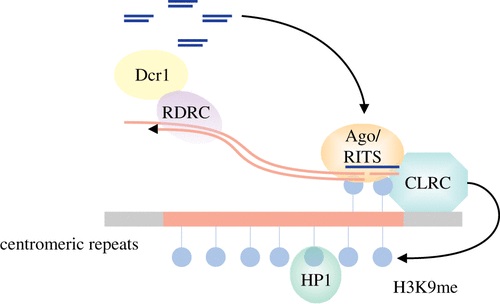
برای مثال در آرابیدوپسیس، متیلاسیون H3 – K9 بطور مستقیم منجر به متیلاسیون DNA می شود. DNA متیله شده می تواند بعدا با پروتئین های متصل کننده متیل، کمپلکس تشکیل دهد. بدنبال اتصال، ساختار کروماتینی به مقدار زیادی فشرده و متراکم می گردد، در نتیجه رونویسی از ژن های آن ناحیه هتروکروماتین متوقف می گردد.
در این روش، در اولین مرحله، RNA پلیمراز IV بطور مستقیم یک RNA تک رشته ای تولید می کند که این RNA تک رشته ای به RNA دو رشته ای (dsRNA) تبدیل می شود، در ادامه آنزیمی شبیه دایسر ، RNAهای دو رشته ای را به RNAهای تک رشته ای ۲۶ نوکلئوتیدی (siRNA) برش می دهد. در ادامه RNAهای تک رشته ای تولید شده توسط آنزیم شبه دایسر در یک کمپلکس مورد استفاده قرار می گیرند. این امر منجر به تشکیل یک کمپلکس راهنما می شود و این کمپلکس سبب اتصال آنزیم متیله کننده DNA به ناحیه هدف می گردد. همچنین مشخص شده است که افزایش متیلاسیون DNA احتمالا تشکیل هتروکروماتین را القا می کند. متیلاسیون DNA، اتصالات پروتئینی را که در شناسایی سیتوزین متیله شده نقش دارد، افزایش می دهد که منجر به باز آرایی مجدد کروماتین می شود و از این طریق از اتصال عوامل رونویسی جلو گیری می کند